軸索輸送による神経細胞内の情報伝搬機構とその神経変性疾患への関与
背 景(シグナリングエンドソーム仮説)
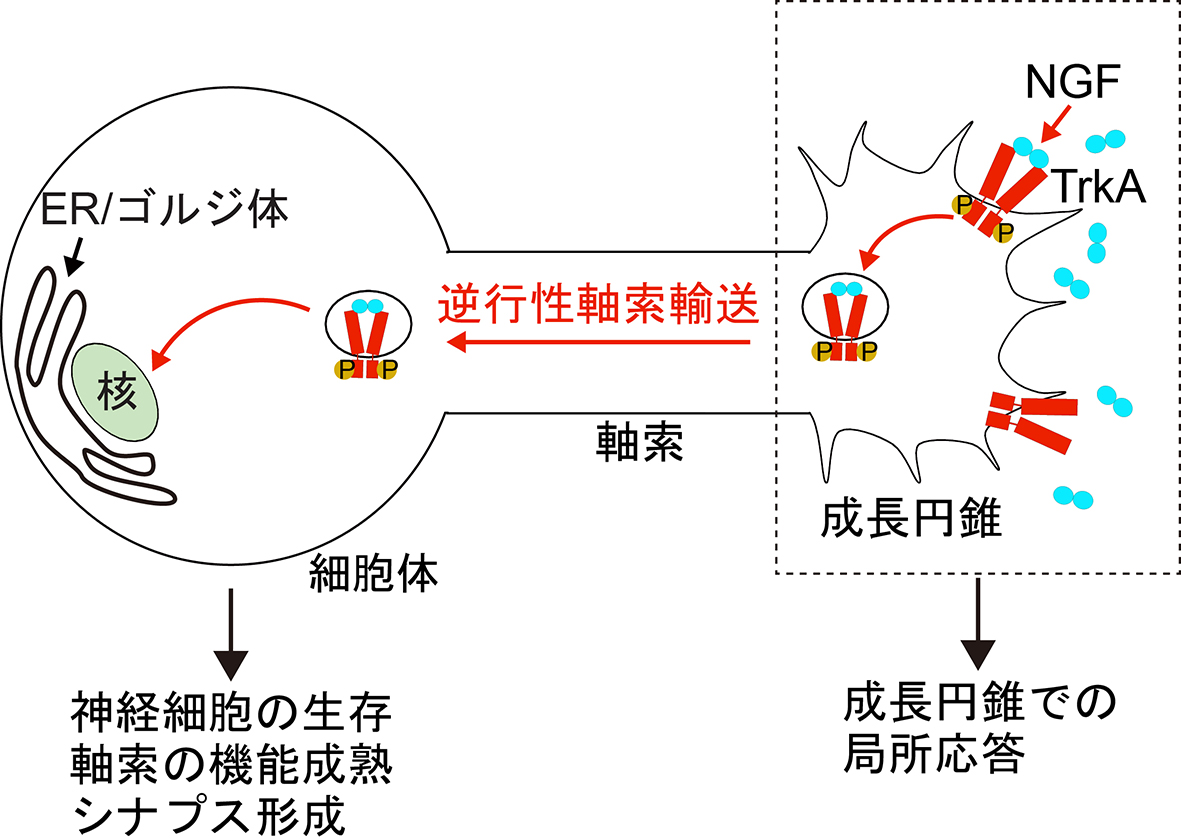
神経細胞は、細胞体、樹状突起、軸索といった細胞区画を有する高度に極性化した特殊な細胞です。この中で軸索は、樹状突起が受容した情報を出力する役割を持つ細胞区画です。しかしながら、他の細胞区画と比べて非常に長い突起であり、軸索先端(成長円錐)で受容した細胞外シグナルを、遠く離れた細胞体に伝える役割も持っています。このように、通常とは「逆方向」に情報を伝搬する機構は「逆行性シグナル」と呼ばれています。逆行性シグナルの実態については未解明な点が多いですが、神経栄養因子の一つである神経成長因子(NGF)をモデルとして研究がなされています。NGFは軸索の標的細胞から分泌され、成長円錐に局在する受容体(TrkA)を介し、標的細胞まで正確に投射した神経細胞を生存、成熟させます。この時、標的細胞に投射したことを細胞体に知らせるため、成長円錐で形成されたNGF/TrkA複合体は、エンドサイトーシスにより取り込まれ、形成されたエンドソームが細胞体方向へ逆行性軸索輸送されることが明らかにされています(図1、文献1)。この機構は、エンドソームを介してシグナルを伝えることから、「シグナリングエンドソーム」と呼ばれています。
|
| 図1.シグナリングエンドソームによる逆行性シグナル伝搬 NGFは成長円錐に局在するTrkAに結合し、リン酸化を介してTrkAを活性化します。NGF-TrkAシグナルは成長円錐局所の応答を誘導するだけでなく、エンドサイトーシスにより成長円錐内に取り込まれた後、細胞体まで逆行性軸索輸送されることで、細胞体まで情報を伝えています。 |
Sema3Aシグナリングエンドソームの発見
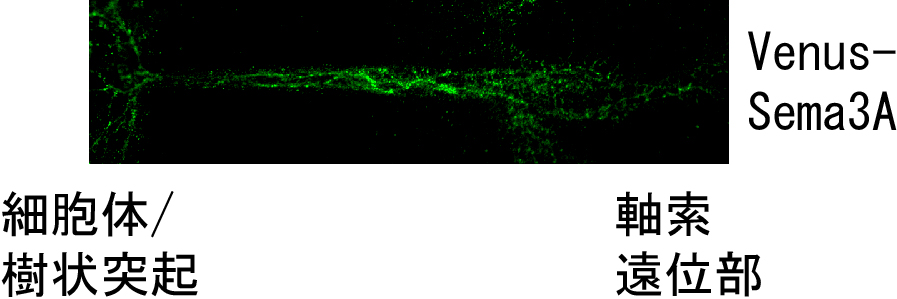
これまで、シグナリングエンドソームによる逆行性シグナルの存在は、NGF等の神経栄養因子でのみ明らかにされており、他の種類の細胞外シグナルにおいても存在するかは不明でした。我々は、セマフォリン3A (Sema3A)という、軸索の伸長を抑制する分子が、シグナリングエンドソームを介した逆行性シグナルを惹起することを発見しました。Sema3Aは軸索先端の成長円錐に作用する分子として発見されましたが、軸索のみならず樹状突起にも作用することが明らかにされています。そこで、一つの細胞外分子が、どのように軸索と樹状突起の両方に作用するかを研究したところ、Sema3Aの作用点が軸索の成長円錐に限局し、軸索の伸長抑制と樹状突起成熟を同時に制御することが分かりました。この時、成長円錐で形成されたSema3Aとその受容体Neuropilin-1、PlexinA4を含む複合体は、成長円錐内にエンドソームとして取り込まれた後、細胞体、さらには樹状突起まで輸送されることで樹状突起にも作用することが明らかになりました(図2、文献2)。 興味深いことに、Sema3Aシグナリングエンドソームは、Sema3A受容体の一つであるPlexinA4と、NGF受容体のTrkAとの相互作用を介して、NGFシグナリングエンドソームとクロストークすることが分かりました(文献3)。そこで現在は、このクロストークによるSema3A/NGFシグナルのバランス制御が、神経回路形成や神経変性疾患発症を制御する分子機構を研究しています。
|
| 図2.Sema3Aシグナリングエンドソームの発見 Venus-Sema3Aを軸索遠位部に局所添加すると、細胞体・樹状突起まで輸送されることから、Sema3Aシグナリングエンドソームの存在が明らかになりました。 |
NGFシグナリングエンドソームとトランスサイトーシス
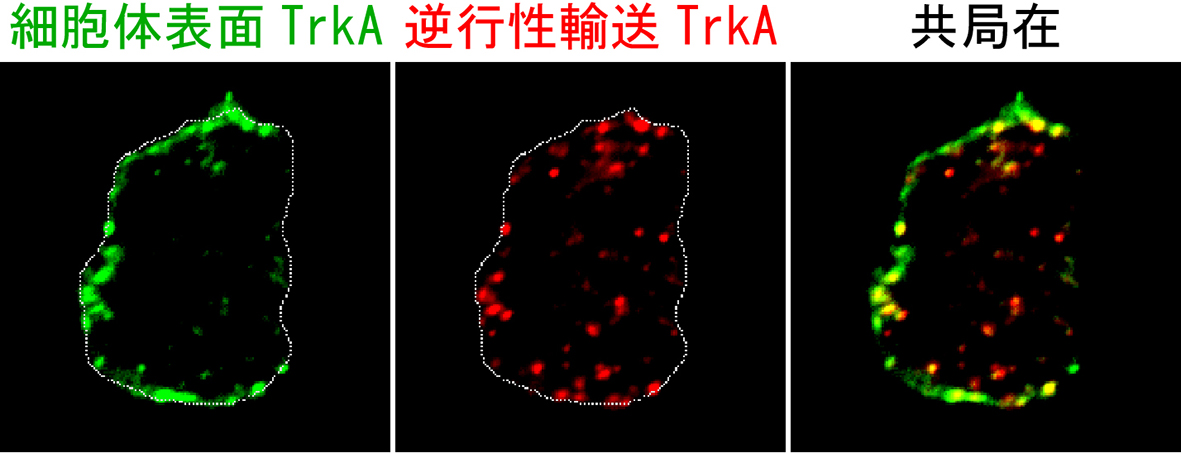
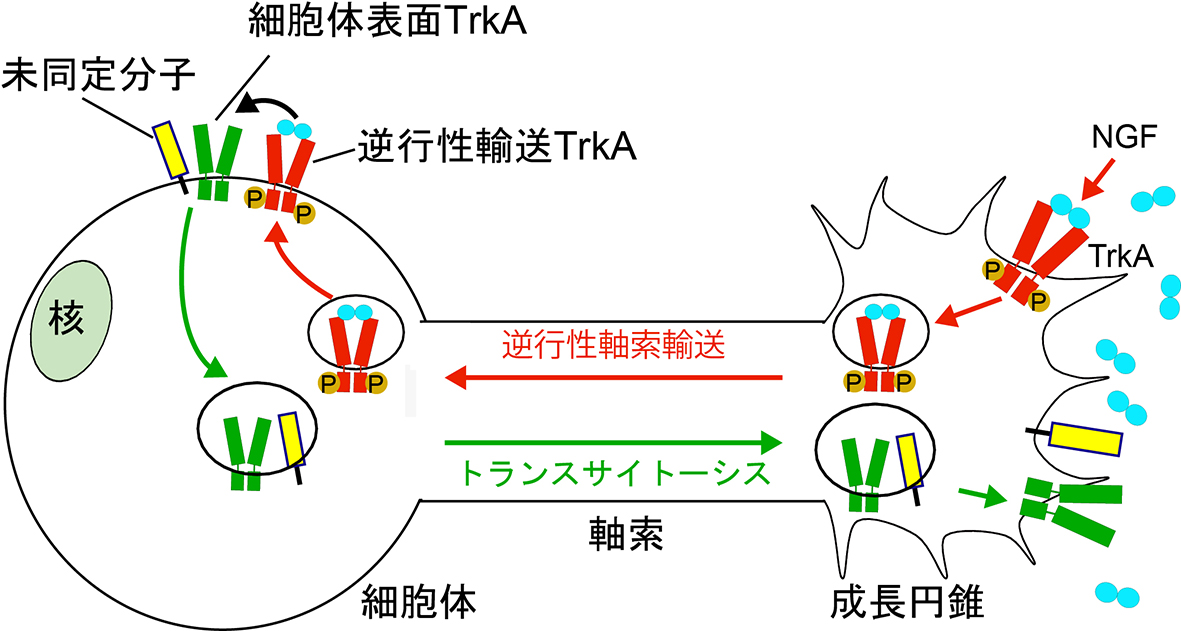
Sema3Aシグナリングエンドソームの発見や、Sema3A、NGFシグナリングエンドソームのクロストークを発見したことから、NGFシグナリングエンドソームの研究にも着手しました。これまで、NGFシグナリングエンドソームは、軸索から細胞体まで輸送された後、核に作用し、種々の遺伝子発現を調節する作用があると考えられてきました。しかしながら、Sema3Aシグナリングエンドソームと同様に、樹状突起まで輸送されることが明らかにされるなど、これまで考えられてきたよりも多様な役割があることが示唆されています。我々は、細胞体まで輸送されてきたTrkA(逆行性輸送TrkA)が、細胞体膜に表出し、細胞体膜に局在していたTrkA(細胞体表面TrkA)と相互作用することを見出しました(図3)。相互作用することで、シグナリングエンドソームを介した情報を受け取った細胞体膜由来TrkAは、その後、細胞体内にエンドサイトーシスされてから、成長円錐まで軸索輸送されました。この機構は、成長円錐でNGFを受容できた(正確に投射した)神経細胞が、成長円錐におけるTrkAの局在量を増やすことで、NGFに反応しやすくなるポジティブフィードバック機構であると考えられます(文献4)。
|
| 図3.受容体の相互作用を介した細胞体と軸索のコミュニケーション 軸索から細胞体へ逆行性輸送されたTrkAを赤、細胞体表面由来TrkAを緑で標識した結果、これらが細胞体膜近傍で共局在することが分かりました。 |
細胞体膜上に局在するタンパク質を、成長円錐などの別の細胞区画の膜上に提示する機構は「トランスサイトーシス」と呼ばれています。しかしながら、神経細胞においてトランスサイトーシスされる分子はTrkAを含め4分子ほどしか同定されておらず、神経細胞の発生・成熟・機能における役割はほとんど分かっていません。そこで現在は、NGFシグナリングエンドソームを介してトランスサイトーシスされる分子のスクリーニングなどを通し、トランスサイトーシスを介したNGFの機能発現の詳細な分子機構の解明や、トランスサイトーシスの破綻と神経変性疾患との関係について研究しています。
|
| 図4.トランスサイトーシスを介したNGFの機能発現 軸索成長円錐へのNGF刺激が、TrkAを細胞体から成長円錐へトランスサイトーシスされることが分かりました。そこで現在は、NGF刺激を介してトランスサイトーシスされる分子が他にないかを調べることで、NGFによる軸索の作用が、トランスサイトーシスされる分子群によって制御されるかを研究しています。 |
実験手法
Compartmentalized chamber(図5)や、microfluidic chamberを用い、神経細胞の細胞体・樹状突起と軸索を異なる環境で培養する手法を用い、例えば、軸索領域にのみSema3AやNGFを添加するといった、生体内と類似した環境を再現して実験を行っています。培養した神経細胞は、抗体などを用いて局所標識した標的分子の挙動解析や、タイムラプスイメージング、さらには、タンパク質の発現解析や、リン酸化などの翻訳後修飾解析といった生化学解析に用いることが出来ます。これまでは、実験動物から採取した神経細胞を用いて研究してきましたが、iPS細胞を利用することで、ヒト細胞由来の神経細胞における実験系の構築も進めています。
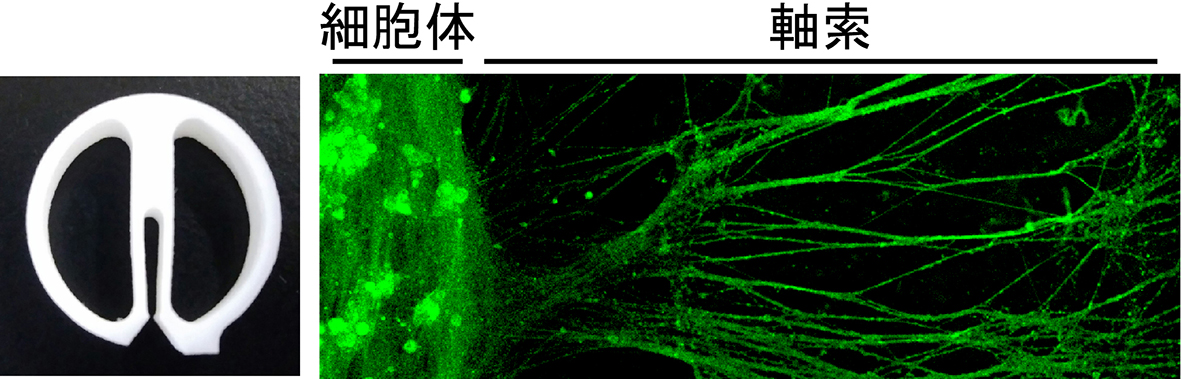
|
| 図5.Compartmentalized chamber Compartmentalized chamber(左図)と呼ばれる特殊な培養チャンバーの中央部分を用いて神経細胞を培養すると、軸索のみが両側の区画まで伸長し、細胞体や樹状突起と分離できます(右図)。中央の区画と両側の区画において培養液がまじりあうことは無く、細胞体・樹状突起と軸索を異なる細胞外環境で培養することが出来ます。 |
参考文献
- Yamashita N and Kuruvilla R.: Neurotrophin signaling endosomes: biogenesis, regulation, and functions. Curr Opin Neurobiol. Vol. 39:139-45, 2016
- Yamashita N, Usui H, Nakamura F, Chen S, Sasaki Y, Hida T, Suto F, Taniguchi M, Takei K, Goshima Y.: Plexin-A4-dependent retrograde semaphorin 3A signalling regulates the dendritic localization of GluA2-containing AMPA receptors. Nat Commun. doi: 10.1038/ncomms4424, 2014,
- Yamashita N*, Yamane M, Suto F, Goshima Y*: TrkA mediates retrograde semaphorin 3A signaling through plexin A4 to regulate dendritic branching. J Cell Sci. Vol 129: 1802-1814, 2016 *Corresponding author
- Yamashita N, Joshi R, Zhang S, Zhang Z.Y, Kuruvilla R.: Phospho-regulation of soma-to-axon transcytosis of neurotrophin receptors. Dev Cell. Vol., 42(6):626-639, 2017